

1. Study of cytokinins
Cytokinins are a class of phytohormones, which enhances cytokinesis of plant cells in the presence of auxin. Over the past half-century, cytokinins have been found to be involved in various aspects of the developmental processes, including senescence, apical dominance, root proliferation, phyllotaxis, and reproductive competence. To regulate the processes, cytokinin metabolism must be tightly controlled. However, the whole picture of the cytokinin metabolic pathway has not yet been completely elucidated. The aim of our research is to fully understand cytokinin functions by elucidating cytokinin metabolism.
TOPICS
- 1-1. Identification of responsible genes for the initial reaction of cytokinin biosynthesis in Arabidopsis.
- 1-2. Identification of enzymes, CYP735A1 and CYP735A2, responsible for the production of trans-zeatin type cytokinins
- 1-3. Discovery of a novel cytokinin-activation pathway
- 1-4. Cytokinin is a key factor regulating the productivity of rice
- 1-5. Cytokinin synthesis by Agrobacterium tumefaciens
- 1-6. Identification of a key enzyme reaction mechanism on cytokinin synthesis based on structural biology
1-1. Identification of responsible genes for the initial reaction of cytokinin biosynthesis in Arabidopsis.
Cytokinin biosynthesis is a key step regulating cytokinin levels in plants. The initial step of cytokinin biosynthesis is catalyzed by adenosine phosphate-isopentenyltransferase (IPT), which uses ATP/ADP/AMP and DMAPP as substrates to generate N6-(��2-isopentenyl)adenine ribotide (iPRPs; Fig.). We have identified a gene family (AtIPT1, AtIPT3-AtIPT8) encoding IPT in Arabidopsis (Takei et al. 2001, JBC).
Nitrogen nutrients are known to increase the cytokinin content in plant roots and xylem sap. We found that regulation of IPT expression by nitrate and/or glutamine-related signals is responsible for the phenomenon(Takei et al. 2004, PCP),(Nobusada et al. 2013, PCP), indicating that nitrogen-responsive cytokinins that burst in the root is regulated at the initial step of biosynthesis. These results support the role of cytokinin in long-distance nitrogen signaling (Hirose et al. 2008),(Kudo et al. 2010).
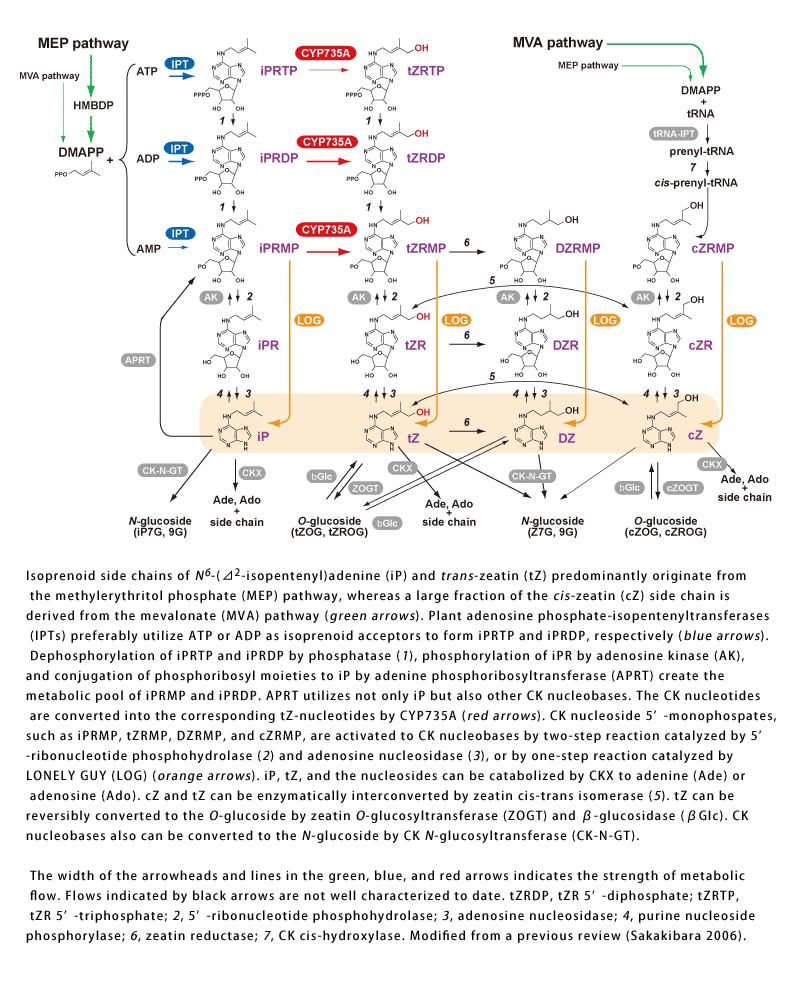
- Figure1-1-1. Current model of isoprenoid cytokinin (CK) biosynthesis pathways in Arabidopsis.
- Isoprenoid side chains of N6-(��2-isopentenyl)adenine (iP) and trans-zeatin (tZ) predominantly originate from the methylerythritol phosphate (MEP) pathway, whereas a large fraction of the cis-zeatin (cZ) side chain is derived from the mevalonate (MVA) pathway (green arrows). Plant adenosine phosphate-isopentenyltransferases (IPTs) preferably utilize ATP or ADP as isoprenoid acceptors to form iPRTP and iPRDP, respectively (blue arrows). Dephosphorylation of iPRTP and iPRDP by phosphatase (1), phosphorylation of iPR by adenosine kinase (AK), and conjugation of phosphoribosyl moieties to iP by adenine phosphoribosyltransferase (APRT) create the metabolic pool of iPRMP and iPRDP. APRT utilizes not only iP but also other CK nucleobases. The CK nucleotides are converted into the corresponding tZ-nucleotides by CYP735A (red arrows). CK nucleoside 5�f-monophospates, such as iPRMP, tZRMP, DZRMP, and cZRMP, are activated to CK nucleobases by two-step reaction catalyzed by 5�f-ribonucleotide phosphohydrolase (2) and adenosine nucleosidase (3), or by one-step reaction catalyzed by LONELY GUY (LOG) (orange arrows). iP, tZ, and the nucleosides can be catabolized by CKX to adenine (Ade) or adenosine (Ado). cZ and tZ can be enzymatically interconverted by zeatin cis-trans isomerase (5). tZ can be reversibly converted to the O-glucoside by zeatin O-glucosyltransferase (ZOGT) and ��-glucosidase (��Glc). CK nucleobases also can be converted to the N-glucoside by CK N-glucosyltransferase (CK-N-GT).
Modified from a previous review (Sakakibara 2006).

1-2. Identification of enzymes, CYP735A1 and CYP735A2, responsible for the production of trans-zeatin type cytokinins
The major forms of cytokinins in Arabidopsis are N6-(��2-isopentenyl)adenine (iP)- and trans-zeatin (tZ)-type cytokinins. Although free-base forms of both iP- and tZ-type cytokinins, iP and tZ respectively, are assumed to be the active forms, several lines of evidence suggest that they have different physiological functions (differential localization of iP- and tZ-type cytokinins in xylem and phloem, and ligand preferences of cytokinin receptors). tZ-type cytokinin is synthesized by trans-hydroxylation of the prenyl side chain of iPRPs (Figure 1-1-1). We have identified two Arabidopsis P450s, CYP735A1 and CYP735A2, which catalyze the hydroxylation step (Takei et al. 2004, JBC). Study on the gene function revealed that physiological role of cytokinins is modulated through side-chain modification (trans-hydroxylation): tZ is required for shoot growth but dispensable for root growth (Kiba et al. 2013). We propose that the control of cytokinin function by side-chain modification is crucial for shoot growth regulation in plants.
3
1-3. Discovery of a novel cytokinin-activation pathway
Free-base cytokinins, which are bioactive forms, had been considered to be generated solely by a two-step reaction; dephosphorylation of ribotides and deribosylation of ribosides. In collaboration with Prof. Kyozuka et al. (Univ. Tokyo), we have found a rice gene LONELY GUY (LOG) encoding phosphoribohydrolase, which catalyzes the direct conversion of ribotides to free-base cytokinins in rice (Kurakawa et al. 2007). In a rice LOG mutant, floral meristem flattens and organ differentiation terminates prematurely without forming the normal number of floral organs . Such genetic evidence and the expression profile of LOG in floral meristem suggest that LOG plays an important role in meristem maintenance at the reproductive stage of rice. We have also characterized LOG family genes in Arabidopsis with multiple knock-out plants and showed that the LOG pathway plays a central role of cytokinin-activation step in plants. (Kuroha et al. 2009),(Tokunaga et al. 2012).
1-4. Cytokinin is a key factor regulating the productivity of rice
The grain number per panicle is an important trait of agricultural productivity in rice. In collaboration with Prof. Ashikari (Nagoya Univ.) and Prof. Matsuoka (Nagoya Univ.) et al., a quantitative trait loci (QTL) analysis was performed using inbred lines derived from a cross between indica cv. Habataki (Ha) and japonica cv. Koshihikari (Ko) to detect QTLs for grain numbers (Ashikari et al. 2005). A Ha plant produces more grains in the main stem than a Ko plant (Figure 1-4-1). The QTL analysis detected 5 chromosomal regions involved in the regulation of grain numbers (Gn1 to 5). Among the QTLs, Gn1a loci has been cloned and shown to encode a cytokinin oxidase/dehydrogenase, OsCKX2. These data show that cytokinin is one of the major determinants of grain yield in rice.

- Figure1-4-1. A panicle of Koshihikari (left) and Habataki (right)
- Habataki produces twice the grain number of Koshihikari. One of the causes for this difference is cytokinin oxidase/dehydrogenase, OsCKX2.
1-5. Cytokinin synthesis by Agrobacterium tumefaciens
Cytokinin is also synthesized by phytopathogenic bacteria, such as Agrobacterium tumefaciens, and is a key factor in some plant diseases. A. tumefaciens infects plants and induces the formation of tumors by integrating the transferred-DNA (T-DNA) region of the Ti-plasmid into the plant nuclear genome. Tumors are formed because the T-DNA encodes enzymes that modify the synthesis of two plant growth hormones, auxin and cytokinin. We had demonstrated that a cytokinin biosynthesis enzyme, Tmr, which is encoded by the Agrobacterium T-DNA region, is targeted to plastids of infected plant cells, and increases cytokinin production by modifying the biosynthetic pathway in the host plant�iSakakibara et al. 2005),(Ueda et al. 2012).
1-6. Identification of a key enzyme reaction mechanism on cytokinin synthesis based on structural biology
Although there is a long history of cytokinin research from biological and agronomic perspectives that has provided a wealth of knowledge about plant development and plant-microbe interactions, understanding the structural basis for the reaction mechanism, substrate specificity, and evolution of cytokinin biosynthesis has been lacking. We have determined crystal structures of Tzs from A. tumefaciens, complexed with a natural substrate or its analogue (Sugawara et al. 2008). The structures provide a structural insight into the reaction mechanism and unique evolution of cytokinin biosynthesis. A structure-based mutagenesis study of Tzs was used to identify the crucial amino acid residues that determine prenyl-donor substrate specificity differences between Tzs and higher-plant IPTs.
2. Hormone analysis platform
Plant hormones play an important role as signaling molecules in the regulation of almost all phases of plant development, from embryogenesis to senescence. Current understanding of hormone action is based on the premise that the signaling systems build a network and mutually regulate signaling and metabolic systems. In order to better understand the network regulation of hormone action, the omics approach, which simultaneously analyzes multiple hormone contents, is clearly important. In our research group, we have developed a highly sensitive and high-throughput method for the simultaneous analysis of more than 40 molecular species of cytokinins, auxins, abscisic acid, gibberellins, jasmonic acid, and salicylic acid including their active forms and derivatives using cutting-edge mass spectrometers. Our current method needs around 10 to 100 mg of fresh weight plant tissues to determine phytohormone profiles and enables us to analyze simultaneously more than 190 plant samples. Using this platform, we are conducting plant hormone research including a number of collaboration studies. Please check our paper for further details of our analysis platform (Kojima et al.2009).
Auto Solid Extraction devices


Analysis devices


